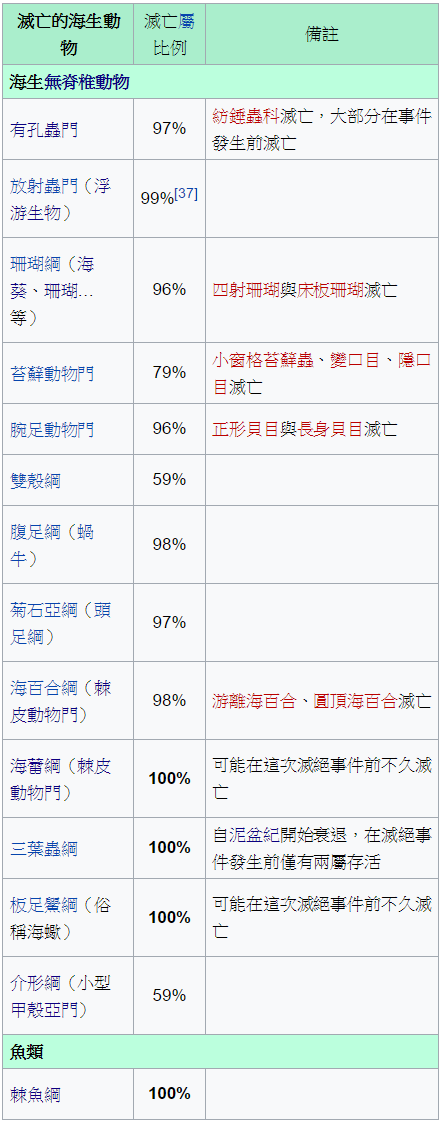
就當然有肉食性合弓綱

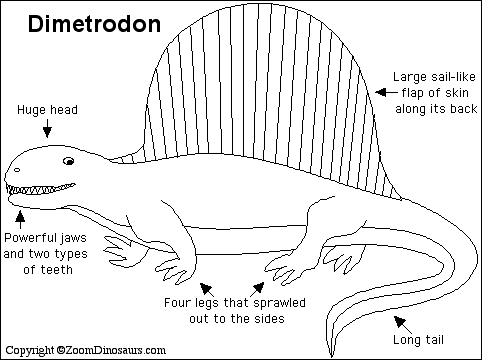
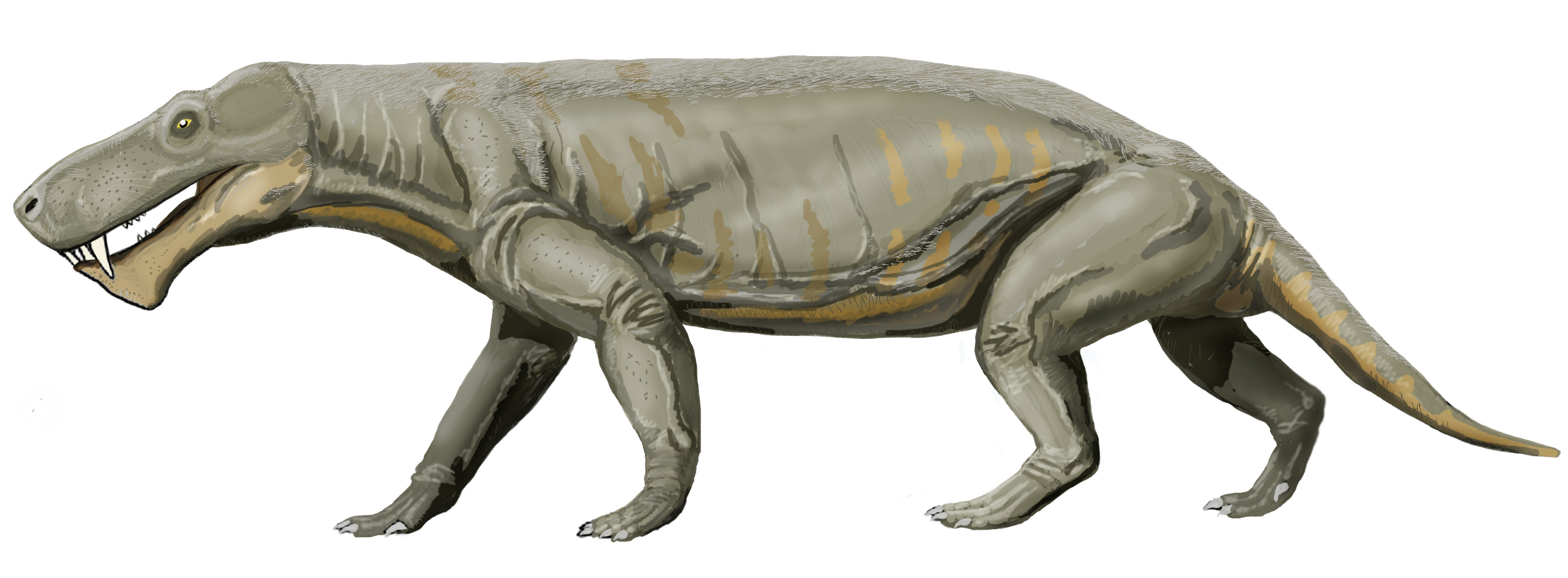
異齒龍 Dimetrodon (= "two measures of teeth")
二疊紀早期,2億9500萬年前 ~ 2億7200萬年前


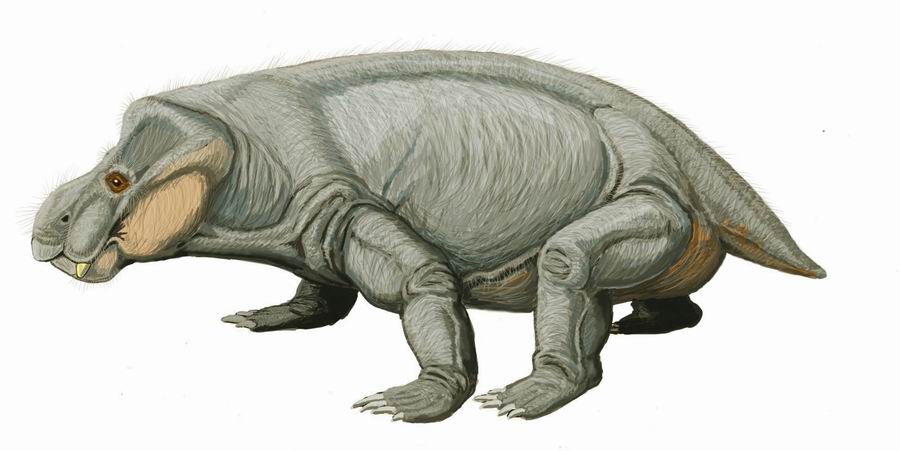
異齒龍(屬名:Dimetrodon)又名異齒獸、長棘龍、兩異齒龍,是肉食性合弓動物(似哺乳爬行動物)的一屬,生存於二疊紀(阿丁斯克階到卡匹敦階),約2億9900萬-2億100萬年前。異齒龍與哺乳類的關係較接近,離真爬行動物(如恐龍、蜥蜴、鳥等)較遠。 (按︰梗係啦,佢係合弓綱,唔係蜥形綱)

儘管一般大眾將異齒龍聯想是恐龍的一份子,但異齒龍其實並不是恐龍。更確切地說,牠們被歸類為盤龍目。異齒龍的化石在北美與歐洲等地均有發現。二疊紀時北美與歐洲的氣候大概像大陸性氣候一樣乾燥,所以異齒龍有很強的適應能力。


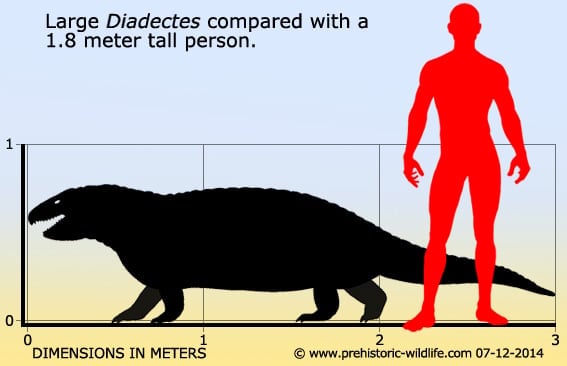
在牠們生存的時代裡,異齒龍是大型頂級掠食動物,身長達3.5米。屬名的意思是「兩種尺寸的牙齒」,因為牠的大型頭顱骨中有兩種不同型態的牙齒(切割用的牙齒與銳利的犬齒)[1]。有這種差異的生物通稱為異齒動物。牠利用往側邊攤開的四肢及大型尾巴來支撐身體。異齒龍也許以類似現今蜥蜴的方式行走。
背帆
異齒龍最明顯的特徵是背部的高大背帆,某些盤龍類(基龍、伊安忒龍、楔齒龍)也有這種特徵。這背帆可能用來控制體溫,背帆的表面可使加熱、冷卻更有效率。這種溫度的調節非常重要,因為可讓牠有更多時間來捕獵獵物。背帆也有可能用作求偶或是嚇阻獵食者。背帆是由脊椎神經棘支撐,每一條連接自個別的脊椎。在1973年,有研究計算一隻200公斤的異齒龍從26℃提升到32℃的體溫,若沒有帆狀物需要205分鐘,但若有則只需80分鐘[2]。





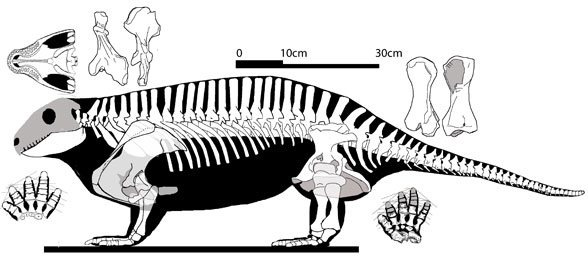





























 在滅絕事件之後,陸地與海洋的生態圈花了數百萬年才完全恢復,比其他大型滅絕事件的恢復時間更長久。此次滅絕事件是地質年代的五次大型滅絕事件中,規模最龐大的一次,
在滅絕事件之後,陸地與海洋的生態圈花了數百萬年才完全恢復,比其他大型滅絕事件的恢復時間更長久。此次滅絕事件是地質年代的五次大型滅絕事件中,規模最龐大的一次,
 某些存活的族系,並未長時間繼續存在,而其他族系則開始多樣化,並長期存活著。有充足證據顯示,超過⅔的陸生兩棲動物、蜥形綱、獸孔目的科在這個滅絕事件中消失。大型的草食性動物遭受嚴重的打擊。除了前稜蜥科,幾乎所有的二疊紀無孔亞綱動物滅亡;龜鱉目具有無孔類的頭骨,被認為可能是從前稜蜥科的近親演化而來(或是雙孔亞綱)。盤龍目早在二疊紀末期前就已滅亡。由於二疊紀的雙孔亞綱化石很少,無法確定滅絕事件對雙孔亞綱所造成的影響;雙孔亞綱包含大部分的爬行動物,例如蜥蜴、蛇、鱷魚、恐龍、以及恐龍的後代鳥類。
某些存活的族系,並未長時間繼續存在,而其他族系則開始多樣化,並長期存活著。有充足證據顯示,超過⅔的陸生兩棲動物、蜥形綱、獸孔目的科在這個滅絕事件中消失。大型的草食性動物遭受嚴重的打擊。除了前稜蜥科,幾乎所有的二疊紀無孔亞綱動物滅亡;龜鱉目具有無孔類的頭骨,被認為可能是從前稜蜥科的近親演化而來(或是雙孔亞綱)。盤龍目早在二疊紀末期前就已滅亡。由於二疊紀的雙孔亞綱化石很少,無法確定滅絕事件對雙孔亞綱所造成的影響;雙孔亞綱包含大部分的爬行動物,例如蜥蜴、蛇、鱷魚、恐龍、以及恐龍的後代鳥類。