到最後,主龍形類(Archosauriformes)演化出主龍類(Archosauria)。
主龍類(Archosauria)
三疊紀早期到現代


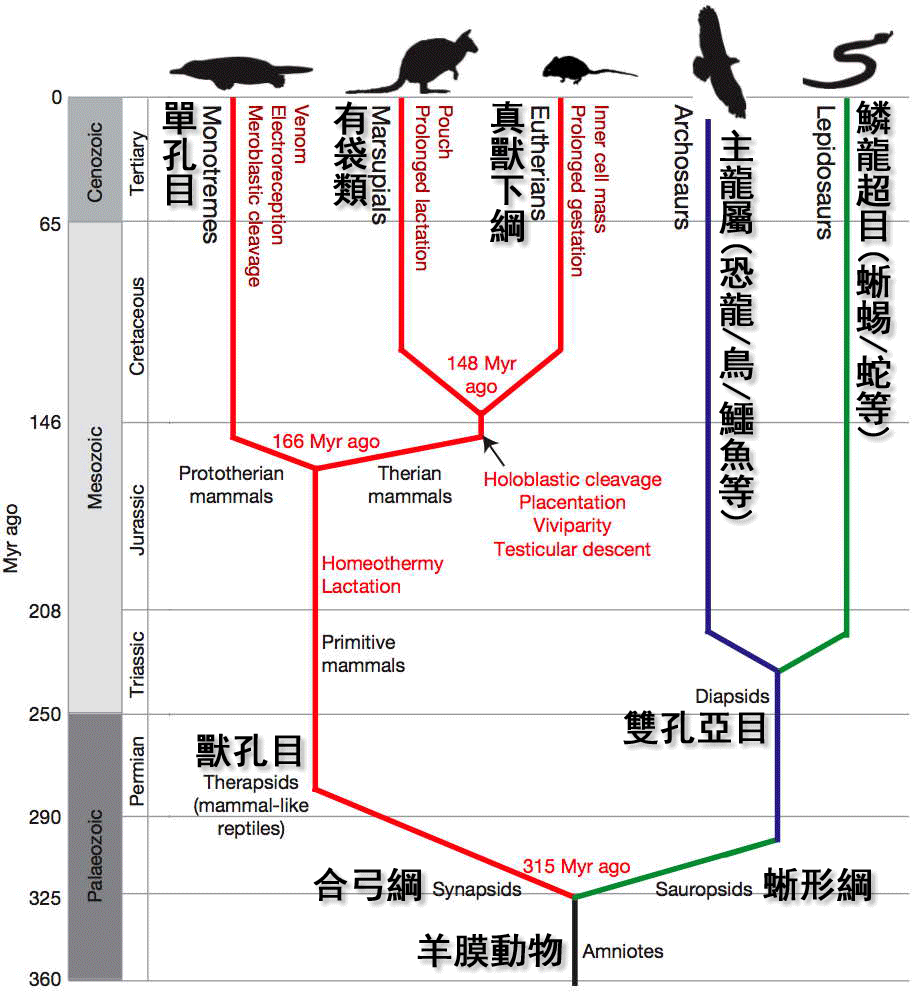
主龍類(Archosauria)又名初龍類、祖龍類、古龍類,在希臘文意為「具優勢的蜥蜴」,是雙孔亞綱爬行動物的一個主要演化支,包含恐龍、翼龍目,現僅存鱷魚與鳥類。
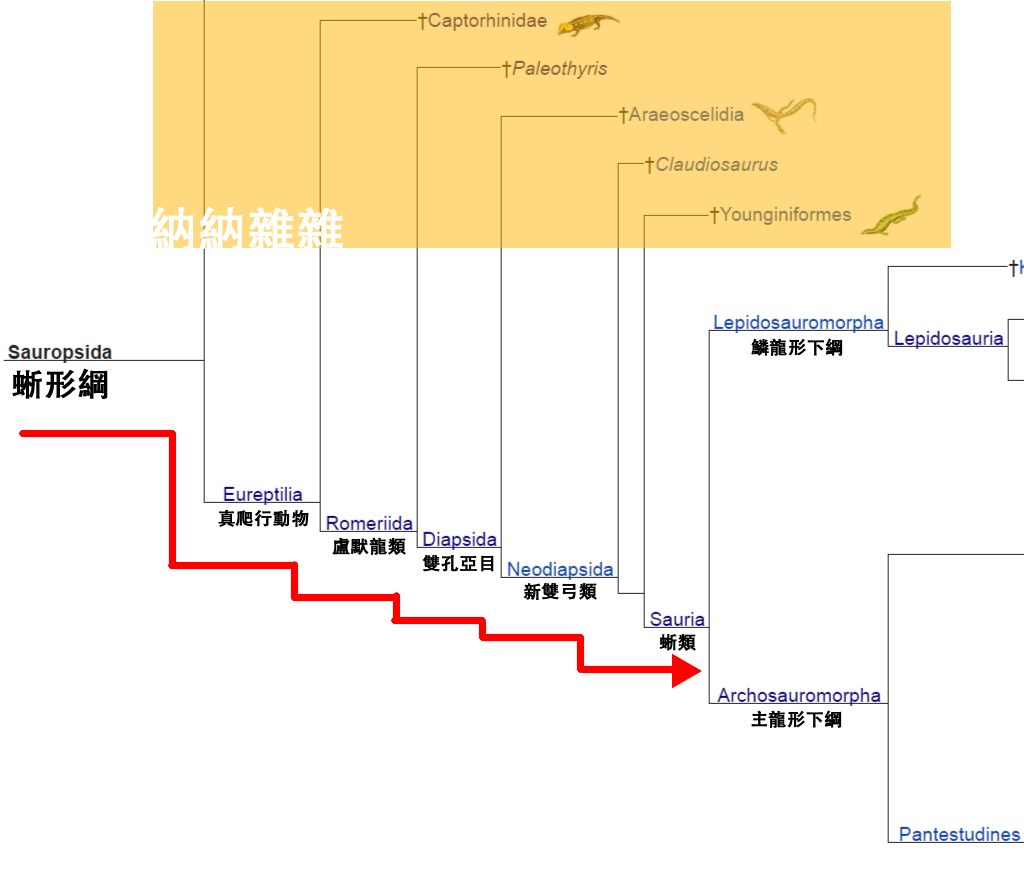
關於主龍類何時開始出現,有許多爭論。有一派將二疊紀的俄羅斯主龍(Archosaurus rossicus)與原龍(Protorosaurus speneri)視為真正的主龍類,這讓主龍類首次出現於晚二疊紀。而另外一派,將俄羅斯主龍與原龍分類於主龍形類(並非主龍類但關係最近),使得主龍類在早三疊紀奧倫尼克階從主龍形類演化而來。
主龍類(Archosauria)在希臘文意為「具優勢的蜥蜴」。但許多中文網站與書籍,誤認學名的意思為「古老的蜥蜴」,而翻譯為初龍類、祖龍類、古龍類。
在某些中文網站中,仍然保持早期的傳統分類法,將主龍類(Archosauria)分類成雙孔亞綱的一個下綱,名為主龍下綱、初龍下綱;或是爬行綱的一個亞綱,名為主龍亞綱、初龍亞綱、祖龍亞綱、古龍亞綱。
主龍類的最基本且最廣為接受的共有衍徵包括:
.牙齒位在齒槽內,這讓牠們進食時牙齒較不易脫落(這也是槽齒目的名稱來源。不過目前槽齒目被視為早期主龍類的並系群集合,不被現代古生物學家所接受)。[img][/img]
.眶前孔(Preorbital fenestrae,位於鼻孔與眼睛之間)減輕了頭顱骨的重量,因為大部分早期主龍類的頭顱骨長而重,這對主龍類是一個非常重要的特徵。眶前孔通常大於眼眶。
.下頜孔(Mandibular fenestrae,下頜骨頭上的小洞孔)稍為減輕了下頜的重量。
.股骨有第四粗隆部(肌肉附著點)。這個特徵似乎並不重要,卻是恐龍演化出直立姿態的關鍵之一(所有早期恐龍以及許多晚期恐龍是二足動物),也可能與主龍類或牠們的祖先在二疊紀-三疊紀滅絕事件中存活下來有關聯。
第1隻既主龍類?
(A) 如果你信第1個說法,"二疊紀的俄羅斯主龍(Archosaurus rossicus)與原龍(Protorosaurus speneri)係真正的主龍類",咁
Archosaurus rossicus 同 Protorosaurus speneri 就係最初個幾種主龍類。
俄羅斯主龍(Archosaurus rossicus)

 原龍(Protorosaurus speneri)
原龍(Protorosaurus speneri)

(B) 如果你信第2個說法,"早三疊紀奧倫尼克階時期既主龍形",咁三疊紀早期既勞氏鱷目,例如Scythosuchus,同Tsylmosuchus,就係最初既主龍類。
勞氏鱷目(Rauisuchia)
(三疊紀早期~晚期)
勞氏鱷目是群所知有限的主龍類集合群,是群生存於三疊紀的掠食動物,大多數體型很大,通常為4到6公尺長。
 呢個不是Scythosuchus或Tsylmosuchus,係勞氏鱷目迅猛鱷屬。
呢個不是Scythosuchus或Tsylmosuchus,係勞氏鱷目迅猛鱷屬。
 勞氏鱷目,係最右邊個種企法。
勞氏鱷目,係最右邊個種企法。
約瑟·波拿巴(José Bonaparte)與麥可·班頓(Michael Benton)先後提出勞氏鱷目,例如蜥鱷,獨自發展出與恐龍不一樣的四肢直立方式;藉由垂直的股骨,與髖臼角度往腹側傾斜,而非股骨頸幹角度傾斜或彎曲。他們將這稱為柱狀直立方式。
直立的步態顯示勞氏鱷類是非常活耀、靈活的掠食動物。勞氏鱷類藉由移動上的優勢,獵食二齒獸類的肯氏獸科。勞氏鱷類是成功的動物,最大的頭顱可達至少1公尺長,直到三疊紀-侏儸紀滅絕事件滅絕了勞氏鱷類與其他大型主龍類。當牠們滅亡後,獸腳類恐龍才有機會成為陸地上的優勢掠食動物;在侏羅紀初期,勞氏鱷類消失,而肉食性恐龍的足跡忽然在尺寸上變大了。







 bio 撚入黎學野
bio 撚入黎學野